无乳链球菌属于β溶血性链球菌,由于该菌细胞壁含有的多糖物质属于Lancefield抗原结构分类中的B族,也称为B族链球菌(GBS)。GBS是寄居在胃肠道和泌尿生殖道的条件致病菌,孕妇的携带率为10%~30%,并且分娩过程中暴露于GBS的新生儿中约2%会出现严重侵袭性感染症状。GBS引发的新生儿肺炎、败血症、脑膜炎等严重感染是世界范围内引起新生儿疾病的主要原因[1]。GBS在成年人中引起的侵袭性疾病也越来越常见,产妇感染可表现为子宫内膜炎,在非妊娠期成年人中可引起皮肤、软组织、骨关节感染和心内膜炎等。
根据其表达的唾液酸荚膜多糖抗原的不同,GBS可分为10种血清型:Ⅰa、Ⅰb、Ⅱ~Ⅸ型。Ⅰ~Ⅴ型最常见,也是定植株最常见血清型。Ⅰa型在产妇GBS相关疾病中最常见[2]。不同血清型与新生儿侵入性疾病的关联性存在差异,Ⅲ型最常见,Ⅰa、Ⅰb、Ⅱ、Ⅴ型次之[2]。Ⅰa、Ⅱb、Ⅴ型在非妊娠期成年人中引起的侵袭性感染中最常见[3]。青霉素是围生期GBS感染的首选治疗药物,为了防止青霉素引起的严重过敏反应,大环内酯类如红霉素、林可霉素类如克林霉素、喹诺酮类和氨基糖苷类抗菌药物通常作为二线药物应用于临床。随着抗菌药物耐药率的增加,疗效也有一定的局限性,进而启用最后一道防线万古霉素,但目前已有万古霉素耐药的GBS分离株的报道。本文旨在综合分析GBS对抗菌药物耐药情况以及概述GBS的耐药机制。
1 青霉素类
青霉素与其他含β-内酰胺环的抗菌药物包括头孢菌素类、碳青霉烯类、单环β-内酰胺类统称为β-内酰胺类抗菌药物。GBS普遍对β-内酰胺类抗菌药物敏感,是治疗GBS感染的一线药物,同样该类药物也可用于围生期GBS感染的治疗。青霉素的β-内酰胺环结合青霉素结合蛋白(PBPs)抑制细菌肽聚糖合成,从而使肽聚糖无法形成交联结构,不能合成完整的细胞壁。同时,不断合成的肽聚糖前体分子触发了水解酶和自溶酶活性,导致细菌裂解。
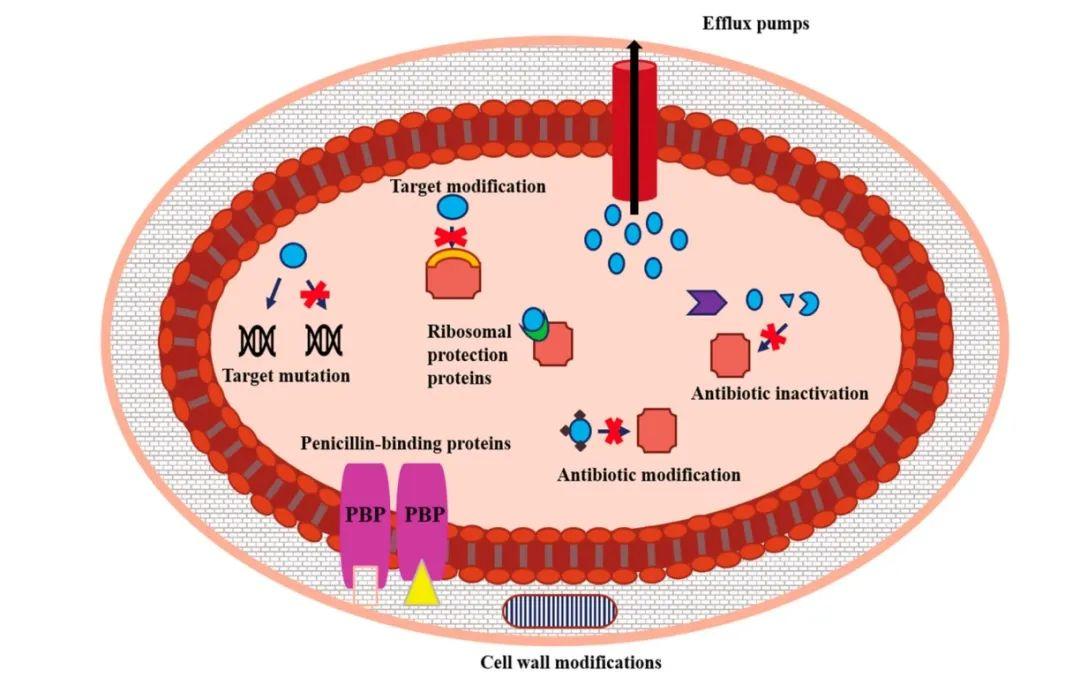
革兰阳性球菌对β-内酰胺类药物的耐药机制主要是使PBPs结构发生变化。有研究发现,PBPs的氨基酸位点发生替换,降低了与青霉素的结合力,使GBS对β-内酰胺类药物敏感性降低[4]。编码PBPs的基因即pbp1a、pbp2a、pbp2b、pbp2x发生突变使GBS获得对青霉素的耐药性。最常见的氨基酸替换发生在PBP2x转肽酶的V405A和(或)Q557E[5]。
PBPs基因突变会导致青霉素耐药株的快速扩散。有学者报道,GBS对青霉素的非敏感率为3.7%[6]。对青霉素敏感性降低通常伴随对其他种类抗菌药物耐药,表现为对大环内酯类和喹诺酮类药物耐药。最近一项研究表明,青霉素耐药的GBS对头孢噻肟、头孢曲松和左氧氟沙星有较高耐药率,分别为28%、36%、93%[7]。目前仍存在对青霉素敏感但是对其他β-内酰胺类抗菌药物敏感性降低的分离株。日本曾报道了对第3代头孢菌素头孢布坦和头孢替安耐药,但对青霉素敏感的分离株[8]。
2 大环内酯类和林可酰胺类
大环内酯类和林可酰胺类是两类化学结构完全不同的抗菌药物,但是作用方式相似。大环内脂类药物与细菌核糖体50S亚基中的23S rRNA结合,阻断肽通道的转肽酶中心(PTC),抑制新生肽段的输出,导致tRNA肽段的裂解。大环内酯类抗菌药物的作用是基于蛋白合成的选择性抑制而非普遍抑制。某些特定的肽段可通过大环内酯类抗菌药物阻断的通道,从而合成蛋白质。这些通过肽通道的肽段可能在随后翻译的过程中被捕获,形成不完整的蛋白质。当新生肽链到达抗菌药物结合的肽通道时,同时抗菌药物结合的核糖体遇到特定的氨基酸组合,蛋白合成被抑制。这种蛋白合成的选择性抑制,以及不完整的蛋白合成导致不受控的翻译,从而加速细菌死亡[9]。林可霉素的作用方式相似,与细菌核糖体50S亚基中的23S rRNA结合,从而干扰蛋白质的合成,这也解释了林可霉素与大环内酯类药物交叉耐药的机制。林可霉素可干扰核糖体A位点,从而阻止了PTC的tRNA肽段的定位。
大环内酯类和林可酰胺类抗菌药物耐药有多种机制,包括外排泵、核糖体修饰、药物失活等。大环内酯类和林可酰胺类药物有相似的结合位点,其中对一种抗菌药物耐药常伴随对另一种抗菌药物耐药。核糖体修饰与GBS对大环内酯类和林可酰胺类广泛耐药有关,而外排泵和药物失活常局限于对某一特定抗菌药物耐药。erm基因能编码甲基转移酶,使核糖体甲基化,是致病菌对大环内酯类药物耐药的主要机制。由mefA/E基因编码的大环内酯外排泵在GBS中较常见,导致对大环内酯类药物耐药。Mef蛋白是主要易化超家族(MFS)成员,不使用ATP作为能量来源将抗菌药物运送至细胞外,而是利用次级主动运输蛋白[10]。ABC-F蛋白介导对与核糖体50S亚基结合的主要抗菌药物的广谱耐药性,包括大环内酯类药物、林可霉素及链霉素A、B,由mre和Isa基因编码。链球菌表达的磷酸转移酶及酯酶可使大环内酯类药物失活,而林可霉素失活受Inu基因编码的核苷酸转移酶调节。美国的一项研究发现,InuB基因通常与IsaC或IsaE基因共存,表明它们之间存在协同作用[11]。
如果患者对青霉素过敏或一线药物治疗无效,可采用红霉素或克林霉素作为二线药物,需要先进行药敏试验。近年来,耐红霉素和克林霉素的GBS急剧增加。有调查结果显示,孕产妇分离的GBS对红霉素及克林霉素耐药率分别可达48.0%及27.4%[12]。
3 四环素
四环素为抑菌性抗菌药物,通过结合核糖体30S亚单位以及抑制蛋白质的延伸发挥抑菌作用。其由线性融合的四环核(A~D环)构成,为功能结合区,主要结合区域为核糖体16S RNA,与氨酰基-tRNA的结合区重叠。
GBS通过表达四环素外排蛋白(TetK和TetL)及核糖体保护蛋白形成耐药[3]。Tet外排基因表达的膜相关蛋白经四环素转运至细胞外,降低了细胞内抗菌药物浓度,从而保护细胞内的核糖体。tetK和tetL基因分别编码TetK和TetL外排蛋白,通常存在于可传递的质粒中,这些质粒可与细菌染色质结合。TetM和TetO是GBS最常见的核糖体保护蛋白,这些蛋白使四环素离开核糖体,进而氨基酰-tRNA分子能与核糖体A位点结合使蛋白质合成继续。
GBS对四环素的耐药率高达70%[13],有多重耐药因素。四环素耐药基因常存在于接合型转座子,促进了细菌间耐药的转移。GBS成人分离株大多数对四环素耐药,特别是tetO和tetM基因的获得,使得这些耐药株群体扩大。耐药GBS克隆株导致其更易于定植及感染,特别是ST-17超毒性克隆株的播散,可导致新生儿严重感染。
4 喹诺酮类
喹诺酮类是一类广谱抗菌药物,常用于治疗呼吸道或泌尿道感染,通过干扰细菌DNA复制过程中的酶从而阻止细菌的生长。喹诺酮类抗菌药物与DNA连接酶或拓扑异构酶结合,导致酶构象发生变化,抑制了酶的正常活性,从而抑制细菌DNA合成,最终导致细胞死亡。
GBS对喹诺酮类药物耐药归因于编码外排蛋白或拓扑异构酶的基因突变。这些突变区域称为喹诺酮耐药决定区,包括编码DNA连接酶的gyrA和gyrC基因,编码Ⅳ型拓扑异构酶的parC和parE基因。这些突变影响喹诺酮类抗菌药物的结合,从而降低抗菌药物的有效性。大多数耐药与喹诺酮耐药决定区基因突变有关,gyrA基因突变常见于丝氨酸-81-亮氨酸(S81L)位点替换,parC常见的基因突变是丝氨酸-79-苯丙氨酸(S79K)位点替换,目前为止仅报道了1例喹诺酮类药物耐药与外排机制相关,且这种外排机制仅仅导致对诺氟沙星耐药[14]。
2003年日本报道了对喹诺酮类耐药的GBS,自此,GBS对喹诺酮类药物的耐药率逐年升高。我国的耐药率也在增加,有研究报道喹诺酮耐药率高达65%[15],显著高于国外,这些分离株多对红霉素、克林霉素、四环素交叉耐药。非妊娠期成年人GBS侵入性感染分离株耐药率可达2.3%[3],CC1克隆株对喹诺酮类药物耐药率高达12.4%[16]。
5 氨基糖苷类和糖肽类
庆大霉素具有杀菌作用,与核糖体30S亚单位的16S RNA的A位点结合从而抑制蛋白质合成。A位点构象发生改变,引起错误的多肽链释放,这些多肽链会损伤细菌的细胞膜。万古霉素通过结合肽聚糖前体的D-丙氨酸-D-丙氨酸末端,抑制肽聚糖的交联。
由于革兰阳性球菌细胞壁对大分子的氨基糖苷类药物渗透性较低,所以GBS本身对庆大霉素低水平耐药。当糖肽靶点氨基糖苷类钝化酶6′-乙酰转移酶-2″-磷酸转移酶发生改变,出现了高水平庆大霉素耐药GBS分离株。细菌对万古霉素的耐药机制为糖肽靶点(D-丙氨酸-D-丙氨酸)的改变,根据被改变的氨基酸分为不同的表型:VanA、VanB、VanD、VanG。其中VanG表型与万古霉素耐药基因vanG(编码D-丙氨酸-D-丝氨酸连接酶)相关,vanG会导致万古霉素结合靶点D-丙氨酸-D-丝氨酸的改变。
万古霉素是治疗GBS感染的最后一道防线[17]。对于一些严重的GBS感染,如感染性心内膜炎、关节置换感染常推荐青霉素和庆大霉素联合治疗。这种联合用药也推荐用于治疗新生儿早发性疾病[18]。但是由于缺乏协同效果研究,以及潜在的肾毒性,这种联合用药治疗方式存在争议。
6 小 结
不同国家、不同地区GBS耐药率的报道存在较大差异,提升对GBS耐药性的检测在GBS的临床治疗中有极为重要的意义。青霉素作为治疗GBS感染的一线药物,通常有效;但是,青霉素敏感性降低的现象越来越普遍。并且二线抗菌药物耐药率较高,新的耐药表型不断被发现。更值得担忧的是,已出现对万古霉素的耐药元件。
GBS对不同种类抗菌药物的耐药性令人担忧,寻找可替代的治疗方法刻不容缓。GBS疫苗正在研发中,但尚未获得使用许可,在此期间需要相应的替代治疗方案。在最近一项试点研究中,妊娠期女性使用靶向益生菌唾液乳杆菌CECT9145可降低GBS阳性率[19]。此外,噬菌体治疗妊娠期GBS感染是一个有效的途径,被证明对降低GBS感染率有极大的益处[20]。天然抗菌药物有望成为抗菌药物的替代品,具有协同作用的联合治疗可减少抗菌药物耐药。GBS临床分离株对乳酸链球菌素有效,红霉素与乳酸链球菌素在治疗GBS中具有协同作用。对这些方案的深入研究有望治疗GBS感染及降低GBS耐药率。
本文荟萃自 《检验医学与临床》2023年20卷7期,只做学术交流学习使用,不做为临床指导,本文观点不代表数字重症 ICU.CN立场。
 微信扫一扫
微信扫一扫